
Abstract
BACKGROUND AND PURPOSE: Deficits in tactile perception are common in patients with PD. However, the neural mechanisms have not been previously reported in the early stages. This study aims to investigate how the brain activity and connectivity changed under tactile perception at early Parkinsonian state by using functional MR imaging.
MATERIALS AND METHODS: Twenty-one patients with early PD and 22 age- and sex-matched controls were recruited and scanned under a passive tactile stimulation task. Within-group and between-group activation maps were acquired, and regions of interest were defined according to the group-comparison result. This was followed by a functional connectivity analysis based on the graph theory.
RESULTS: We found that in the PD group, bilateral sensorimotor cortex was hypoactive during the task, whereas the hyperactive regions were mainly in bilateral prefrontal cortex, bilateral cerebellum, and contralateral striatum. There was a significant decrease of total connectivity degree in ipsilateral SMA in PD, which was negatively correlated with the Unified Parkinson's Disease Rating Scale score. Furthermore, the connection strengths among the areas of prefrontal cortex, striatum, and cerebellum were increased.
CONCLUSIONS: This study illustrated that early PD was associated with not only altered brain activation but also changed functional connectivity in tactile perception. The most significant impairment was in SMA, whereas striato-prefrontal and cerebello-prefrontal loops may play a compensatory role in early PD tactile function.
Abbreviations
- BOLD
- blood oxygen level–dependent
- DLPFC
- dorsal lateral prefrontal cortex
- FWE
- family-wise error
- IFG
- inferior frontal gyrus
- MFG
- middle frontal gyrus
- MNI
- Montreal Neurologic Institute
- PD
- Parkinson disease
- PMC
- premotor cortex
- rCBF
- regional cerebral blood flow
- ROI
- region of interest
- S1
- primary somatosensory cortex
- S2
- secondary somatosensory cortex
- SMA
- supplementary motor area
- UPDRS
- Unified Parkinson's Disease Rating Scale
Modern studies have discarded the traditional perspective that PD is simply a movement disorder. It is now clear that PD is associated with a large number of nonmotor clinical features among which tactile abnormality is a significant part. For example, Schneider et al1,2 reported several tactile deficits in PD, including 2-point discrimination and tactile stimulus location. Similarly, Sathian et al3 revealed that PD patients also were accompanied with deficiencies in tactile spatial acuity and roughness discrimination. Furthermore, both the peripheral nerve and somatosensory-evoked potential recovery curves were normal in patients with PD,4 suggesting these deficits may come from abnormal integration or organization of tactile information in brain level.
It is now deemed that tactile deficits in PD come from disturbed function of the basal ganglia,5 which triggers a series of alterations in the striato-thalamical-cortical loop and finally causes visible changes of activation and connectivity in cerebral cortex.5–8 Such opinion was further supported by several neuroimaging investigations. Weder et al9 found the uptake of 6-[18F]-fluro-l-dopa was significantly reduced in caudate nucleus in PD during tactile discrimination, which was closely related with the diminishment of discrimination function. They also found the rCBF in bilateral sensory and motor cortex was significantly decreased in patients with PD.10 Boecker et al11 reported several hypoactive brain regions in patients under tactile stimulation, such as sensorimotor cortex, prefrontal cortex, basal ganglia, and posterior cingulate. In contrast, ipsilateral S1 and S2 were found hyperactive.
Previous studies have brought us a basic knowledge in Parkinson tactile deficits. First, such deficits did not come from the impairment of the peripheral nervous system but rather are a result of disrupted tactile information processing in the brain.4 Second, such deficits were associated with altered functions in many different brain regions, including basal ganglia, sensory cortex, and motor cortex.9–11 Despite these findings, some problems still remain. Early diagnosis and treatment is very important for patients with PD. Because all the past studies focused on the patients at an advanced or late stage, whether the brain abnormality could be equally found at an early stage was unknown. Furthermore, most of the past studies were simply based on the activation level, but how the altered regions interacted and reorganized in the network level was scarcely known.
Here, we used a tactile task to study brain activations in patients with early PD during tactile perception. We also implemented a connectivity analysis based on graph theory12 to measure the alteration of brain functional connectivity in patients. We aimed to illustrate directly and clearly how the brain function was changed in early PD tactile perception.
Materials and Methods
Subjects
Twenty-one PD patients (10 men, 11 women; mean age, 60.43 years) and 22 healthy volunteers (11 men, 11 women; mean age, 59.23 years) participated in our study. The age and sex differences between 2 groups were tested by 2-sample t test and χ2 test, respectively, and no significances were shown between groups (P > .05). The patients were recruited from the Department of Neurology and the healthy volunteers were recruited from Zhejiang University and the communities. All the subjects gave their written informed consent for the study. The whole procedure was approved by the Medical Ethic Committee of Zhejiang University.
For the PD group, the disease diagnosis was based on the medical history, the neurologic examinations, response to the dopaminergic drugs, scale evaluation, and the exclusion of other neurologic or psychiatric diseases. The patients were asked to pause their drug medications for at least 12 hours before the MR imaging. All the patients were assessed with the UPDRS, the Hoehn and Yahr Scale, and the Mini-Mental State Examination, while off their medications. All the patients were diagnosed at an early stage (H&Y I-II). On-line Table 1 shows the details of the patients.
For both the PD and the control group, we ruled out any subjects who had neurologic or psychiatric disorders, or brain trauma at any time of their lives.
Task Design
We used 2 different tiny wood wheels as the stimuli in the experiment. Both the wheels were the same size with the surface of 1 wheel being smooth and the other wheel being coarse. We carried out the prescan test first. During the test, an operator held the 2 stimuli and rolled the wheel across each subject's skin of the right index finger with a certain pressure. The subjects were asked to close their eyes, passively perceive the stimulation, and respond for the type of stimuli. The response showed that all the subjects could correctly discriminate the 2 wheels during the stimulation. Then, the subjects were invited into the scan room. The experiment went with the block design. Similar to the prescan test, the same operator held the stimuli and rolled the wheel across the subject's skin of the right index finger with a certain pressure in a pseudorandom order. Each stimulus lasted for 3 seconds, and each task block included 10 stimuli, which was 30 seconds. Then, a 30-second resting block followed. Both the task block and the resting block were repeated 3 times, with a resting block at the very beginning, which lasted for 60 seconds. The total scan time was 240 seconds. During the whole procedure, the subjects were instructed to keep their eyes closed and relax their bodies, only focusing on discriminating the stimuli. No subjects felt pain or discomfort during the task according to their feedback after the scan.
Image Acquisition
All the scans were performed on the 1.5T Signa MR scanner (GE Healthcare, Milwaukee, Wisconsin) equipped with a birdcage coil. High resolution axial T1- and T2-weighted anatomic images were first obtained to exclude any dormant neurologic diseases. Functional images were acquired by using a gradient recalled-echo echo-planar imaging sequence. The scan parameters were as follows: TR = 3000 ms, TE = 35 ms, flip angle = 90°, FOV = 240 mm × 240 mm, matrix = 64 × 64, section number per volume = 22, section thickness = 5 mm, and section distance = 1 mm.
Data Analysis
All the image data with DICOM format were first converted into files with Analyze format by using MRIcro software (http://www.sph.sc.edu/cond/rorden/mricro.html). We discarded the first 10 time points (30 seconds) for the instability of the initial MR signals and the adaption of the subjects to the circumstances. The remaining images were analyzed with SPM2 software (Wellcome Trust Centre for Neuroimaging, London, United Kingdom) based on the Matlab6.5 platform (MathWorks, Natick, Massachusetts). The preprocessing began with the realignment for the motor correction, and all the subjects with head motion >2 mm or rotation over 1° in x-, y-, and z-axes were discarded. That comprised 7 subjects with 4 patients and 3 controls, leaving 17 patients and 19 controls. Then, the images of each subject were normalized into the MNI template by resampling with 3 mm × 3 mm × 3 mm. After that, all the data were spatially smoothed by a Gaussian filter of 6-mm full width at half maximum to reduce the spatial noise.
The statistical analysis was performed at 2 levels. First, individual task-related activations were evaluated by using a general linear model. The BOLD signals were estimated at each voxel by modeling with a hemodynamic response function. A high-pass frequency filter (128 seconds) was applied, and low-frequency noise was removed. For the second level, we analyzed all the subject-specific contrast images by using a random effect model. One-sample t test was applied to show the brain activation under tactile stimulation within each group. The statistical threshold was set at P < .05, corrected by FWE with activation voxels >10. We used a 2-sample t test to study the activation differences between PD and controls under the task. The statistical threshold was set at P < .005, uncorrected with activation voxels >5. The activated voxel coordinates were transformed from MNI templates into Talairach space13 by using TalairachClient,14 and all the activated brain regions were presented.
We used MarsBar toolbox15 for the ROI definition. Both the hyperactive and the hypoactive brain regions shown in the between-group activation map were related to the tactile deficits in PD; thus, they were taken to build ROIs. The peak voxel in each cluster was defined as the center and a sphere with a radius of 6 mm was drawn for each of the clusters. We then excluded all the unactivated voxels in each sphere, and the voxels left in the sphere were finally defined as a ROI. The mean BOLD time series of each ROI was extracted from each subject.
In this study, we adopted a network model based on graph theory to describe the functional connectivity (for technical description, see Jiang et al, 200412). All the ROIs were referred to as the “nodes,” and the connections among them were considered as the “links” in the network. The total connectivity degree Γi of a node i in a graph was the sum of all the connectivity degrees between i and other nodes, describing the amount of information received by node i from the particular network. A larger Γ reflected this region had more functional connections with other brain regions and, therefore, played a more important role in the network. We calculated the Γ value of each ROI and used 2-sample t test to compare the differences in Γ between the 2 groups. The statistical threshold was held at P < .05. For those nodes with significant Γ change in group comparison, we investigated the relationships between Γ and the UPDRS scores with Pearson correlation analysis, with the aim to find out whether these changes were related to the disease severity. We also measured the temporal correlations of every 2 nodes to investigate the connection strength of each link and the correlation coefficient r was then converted into Fisher z-value. A 2-sample t test also was implemented for the comparison of z-values between groups. To reduce the false-positive results that may be caused by the multiple comparison, we held the threshold at P < .005.
Results
Activation Map within Groups
Within the normal group, bilateral S1 and contralateral anterior cerebellun were strongly activated under the tactile state, whereas in the PD group, the activated regions involved contralateral S1, ispilateral anterior cerebellum, and contralateral precentral gyrus. All of these regions (Fig 1) were closely related to tactile function in human beings.16–19

Within-group activation maps in controls (A) and patients with early PD (B). The hot colors indicate the activated regions under tactile perception (P < .05 with FWE correction, voxels >10).
Activation Map between Groups
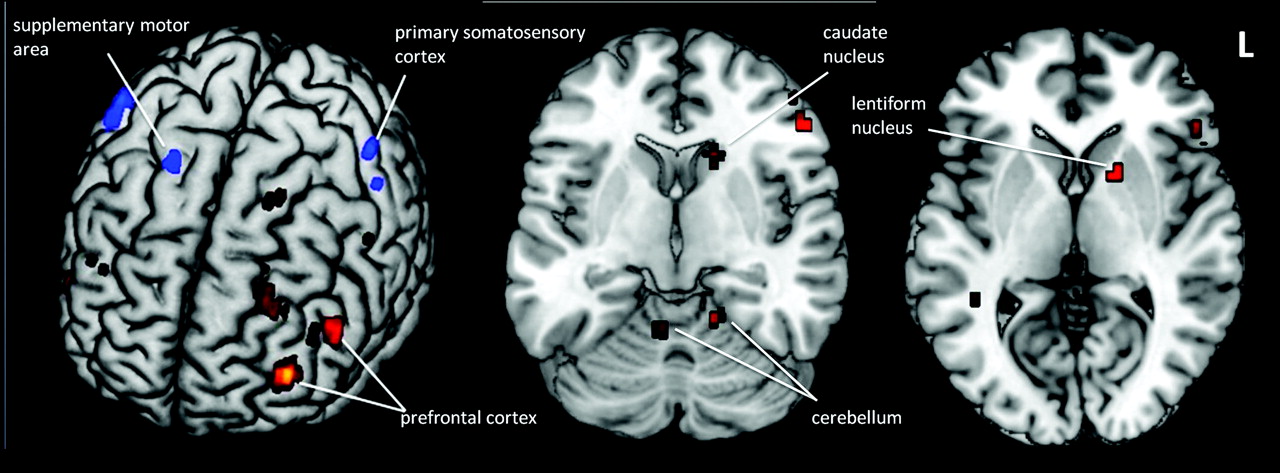
There were several regions showing hypoactive activation under the tactile perception in PD. These included bilateral S1, ipsilateral SMA, ipsilateral PMC, and ipsilateral paracentral lobule. Conversely, many more regions were hyperactive: DLPFC, bilateral anterior cerebellum, bilateral posterior cerebellum, bilateral parahippocampal gyrus, contralateral IFG, contralateral MFG, contralateral caudate nucleus, and contralateral lentiform nucleus (Fig 2 and On-line Tables 2 and 3).

The hyperactive (red) and hypoactive (blue) brain regions in a patient with PD compared with the control under tactile perception (P < .005, uncorrected).
Functional Connectivity between Groups
As a result, all the hypo- and hyperactive regions above were drawn as ROIs. After the functional connectivity analysis, we found there was a significant decline of Γ value in ispilateral SMA in PD patients compared with the controls (P = .032). Furthermore, this value was negatively correlated with UPDRS score (r = −0.57, P < .01; Fig 3)

Correlation between connectivity degrees (Γ) and UPDRS scores in ispilateral supplementary motor area in PD.
In terms of z-value, it decreased in only 1 link, which was between right SMA and right PMC (P = .000) but increased in 4 links: between left IFG and left DLPFC (P = .003), between left DLPFC and right arterior cerebellum (P = .004), between left IFG and left caudate nucleus (P = .001), and between right arterior cerebellum and left lentiform nucleus (P = .002; Fig 4).

Reduced (A) and enhanced (B) connection strengths (z) in brain links in patients with PD compared with controls (P < .005).
Discussion
This research investigated the changes of brain activation and connectivity in early stage of PD during tactile perception. We not only found different activation patterns but also altered functional connectivity in the patients. We noticed that there were no significant differences in performances between the 2 groups. This strongly indicated that the altered brain functions occurred earlier than the tactile symptoms in PD, which should raise an alert. To our knowledge, this is the first research to show the changes of brain functions in early PD tactile perception.
Altered Functions in Sensory and Motor Areas
Functional changes in sensory and motor areas was the first finding in this study. As shown in the results, several brain regions in sensory and motor areas presented a reduced activation. These regions included bilateral S1, ispilateral SMA, contralateral PMC, and ispilateral paracentral lobule. This was consistent with a former study by Weder et al,10 who discovered decreased rCBF in sensorimotor areas at advanced disease state. Our study proved this finding again and further indicated the existence of such deficit even at the early stage. These areas are widely acknowledged to associate with tactile perception and discrimination in the brain,16,20–22 which exactly reflected the tactile deficits in PD.
The most remarkable alteration was in the SMA. Previous studies have revealed that PD was accompanied with hypoactive SMA in several motor tasks, such as finger movement23,24 and joystick movement.25 Here, we also found the SMA was hypoactive under tactile stimulation task, which may indicate that dysfunction of SMA is prevalent in PD, not only related to the motor symptoms but also to the sensory impairment. What is more, apart from activation abnormality, such dysfunction was consistently discovered with functional connectivity analysis. In terms of Γ value, the ispilateral SMA significantly dropped in the PD group compared with the control group. This meant that SMA had less functional connections with other brain regions among the patients and therefore, its importance in the network was declined. Furthermore, its Γ value was negatively correlated with UPDRS scores, indicating the close relationship with the severity of the disease. This certainly reflected a noticeable functional deficit of SMA in early PD that may be an important factor contributing to the tactile impairment.
It was worth noting that the decreased functional connectivity in SMA also has been reported in some other research when patients were under resting state.26,27 Taking this into consideration, SMA really played an important role in PD's neural mechanism, whether in rest or in task. Specific to our study, the reason for the SMA dysfunction in tactile perception could be explained in 2 ways. First, SMA was a significant part in the somatosensory circuitry. It not only exchanged sensory information with subcortical nucleus such as basal ganglia28–31 but also directly connected to somatosensory cortex, such as S1 and S2.28,32 This suggested that the sensory information received from the upper region must undergo a sophisticated processing in the SMA before further transmission to the next station. Thus, the reduced functional connections of SMA in PD seemed to be a consequence of disturbed information input or processing in the tactile perception. Second, it has been well demonstrated that SMA functions in sensorimotor integration,33 which initiates and organizes voluntary movements in line with the sensory stimulation received from the outside.28,34–36 Previous research has shown that attention to somatosensory stimulation was directly linked to the preparation for action, which activated SMA in healthy subjects.37 In accordance with these findings, SMA dysfunction perhaps was also a reflection of impaired sensorimotor integration in PD patients.
We speculated that the decreased Γ value in SMA must be due to some declined z-values in the links between SMA and the other regions. As expected, the link between SMA and PMC dropped. This result may at least partly illustrate where the decreased functional connection of SMA came from. It was found that PMC participated in sensory-guided or sensory-triggered movements,34,38 accepting sensory input from the parietal lobe, and influenced movement through its projections to the motor cortex.39,40 It was also found that the connection between SMA and PMC was decreased in PD when patients were attending to action.41 Therefore, the alteration of the SMA-PMC link possibly reflects a sensorimotor-related deficit in early PD.
Altered Functions in PFC, Striatum, and Cerebellum
Another interesting finding was the altered functions in prefrontal cortex, striatum, and cerebellum under tactile perception in these early patients. These areas, unlike the sensorimotor areas, showed strong enhancements both in the activation and the connectivity. For example, bilateral DLPFC, contralateral IFG, contralateral MFG, bilateral anterior cerebellum, bilateral posterior cerebellum, contralateral caudate nucleus, and contralateral lentiform nucleus were all hyperactive in the tactile task. Moreover, the connection strengths between IFG and DLPFC, between DLPFC and anterior cerebellum, between IFG and caudate nucleus, and between anterior cerebellum and lentiform nucleus were also increased. These “upward” changes, in our opinion, may reflect the compensatory mechanisms in early PD tactile function.
Prefrontal cortex was found to be hypoactive under tactile stimulation in PD in a previous study,11 which seemed to be inconsistent with our results. We speculated this mismatch may owe to the compensatory effect at the early stage. A similar finding has been reported previously, that dopamine uptake was significantly higher in prefrontal cortex in early PD.42 Moreover, hyperactivation of prefrontal cortex was consistently found in patients performing different tasks, such as automatic movement,43 feedback processing,44 and working memories,45 suggesting the potential compensatory mechanisms in PD. As one of the most complicated parts of the brain, prefrontal cortex was recognized for its role in cognitive functions.46 Both the striatum and the cerebellum were closely related to prefrontal cortex, constituting 2 separate loops (striato-prefrontal loop and cerebello-prefrontal loop), which worked together to serve for the higher cognitive works in both humans and nonhuman primates.30,47,48 Besides, the functions of these 2 loops were closely related to dopamine,49–51 which may change in parkinsonian patients. According to these findings, the raised activations and enhanced connections among prefrontal cortex, striatum, and cerebellum in our study probably reflected the alterations of these subcortical-cortical loops, leading to compensations at the early stage. One reasonable interpretation was the patients' drawing on cognitive mechanisms to compensate for their tactile deficits. In other words, they needed more attentions or cognitive efforts to perceive and discriminate the stimuli than the healthy controls did. Yet, this concept needs more studies to support it.
Limitations of the Study
Unavoidably, there were some limitations in this study. First, the tactile test we used in this experiment was relatively simple and could not be quantitatively evaluated. It would be much better to perform a detailed and thorough tactile threshold examination by using neuropsychologic methods to give a full assessment to all the subjects' tactile functions. Second, although all the patients were off medication for at least 12 hours, an influence of the medication cannot be excluded, given the long serum half-time and receptor binding of some dopaminagonists (eg, pergolide, cabergoline, see On-line Table 1). Hence, we should not ignore the potential influences on the results by the medications. Finally, it would be better if a peripheral nervous test was given to exclude any possible peripheral impairment, considering different disease mechanisms may exist among individuals with PD.
Conclusions
In this functional MR imaging study, we not only provided evidence for activation abnormality in early PD tactile function but also demonstrated the functional connectivity changes related to this deficit. Overall, SMA was the most impaired region perceiving tactile stimulation at early-stage PD due to its hypoactivation during the task, declined connectivity degree in the network, and close correlation with the disease severity. Furthermore, this study indicated that the cognitive mechanisms with striato-prefrontal and cerebello-prefrontal loops may play an important role in functional compensation of tactile deficits in patients with early-stage PD.
Acknowledgments
We thank Dr Xuning Zheng (Department of Neurology, First Affiliated Hospital, Zhejiang University School of Medicine) for assistance in patient recruitment. We are also grateful for Drs Huandong Li, Yong Liu, and Tianzi Jiang (Institute of Automation, Chinese Academy of Science) for technical help with data analysis.
Footnotes
-
This work was supported by grant 30570536 from the National Science Foundation of China (to M.Z.) and grant 30900358 from National Science Foundation of China and grant 2009QN005 from the Medical Scientific Research Foundation of Zhejiang Province (both to X.X.).
Indicates open access to non-subscribers at www.ajnr.org
References
- Received January 18, 2011.
- Accepted after revision March 14, 2011.
- © 2011 by American Journal of Neuroradiology




{kind=link}
{kind=link}
{kind=link}
{kind=link}